|
Моделирование самоорганизующихся модулей в биологических системах Томский сельскохозяйственный институт Научный руководитель: Шипилин Николай Николаевич, д.с.-х.н. В последние годы наука о самоорганизации выходит за рамки синергетики и высказывается мнение о необходимости учета геометрических свойств реального пространства [1]. Геометрический аспект, наряду с термодинамическим и информационным, становится одним из основных при моделировании процессов самоорганизации и особенно в биологическом морфогенезе. В отличие от евклидовой геометрии линейных и нелинейных событий эллиптическая геометрия Римана допускает процессы спирально-циклической самоорганизации [2]. Существует система координат в которой замкнутое эллиптическое пространство биобъекта обладает зональным строением. Такое строение пространства, независимо от заполнения его веществом, является заданной возможностью для развития спирально-циклических направленных форм движения. В ранних моделях морфогенеза основное внимание уделялось механизмам становления биохимического паттерна морфофункциональных модулей, а вопросы их пространственной геометрии оставались на втором плане. Зависимость латерального транспорта от общей гауссовской кривизны и другие проявления диссипативных структур позволили сформулировать новый класс моделей, основанный на положении о взаимообусловленности динамики и геометрической формы паттернов модулей. Изучению механизмов взаимообусловленной динамики паттернов модулей от внутренней и внешней их геометрии может способствовать моделирование самоорганизации двумерных (2D) каналов коллективного направленного переноса морфогенов. Предполагается, что необходимым условием морфогенеза является передача информации с помощью волн дифференцировки [3]. Представляется возможным с помощью эллиптической геометрии Римана рассмотреть самоорганизацию модулей дифференцировки с межмодульными 2D каналами, в замкнутом пространстве которых происходит направленный перенос морфогенов и волн дифференцировки.Построение модели и обсуждение В рассматриваемой модели точки являются центрами биоединиц и система точек изучается без учета их природы. В зависимости от масштабного уровня иерархии спирально-циклической самоорганизации такими единицами могут быть клетки и группы клеток. Плоский каркас в евклидовой геометрии заменяется каркасом на поверхности Клиффорда (К-поверхности) в эллиптической геометрии Римана [2]. В соответствии с выбранной симметрией движения самоорганизация модулей дифференцировки ведется поэтапно с помощью биоединиц, находящихся на минимальном расстоянии от нулевой точки. Биоединицы отстоят друг от друга на несколько периодов формируемого каркаса. С помощью системы точек на К -поверхности (К-система) находятся точки расположения биоединиц заданного уровня самоподобия. К-система переносится параллельно самой себе по спирали от периферии к центру и точки фиксируются в пределах треугольного контура от нулевой до n-го масштабного уровня иерархии. Оболочки биоединиц с пониженной размерностью обеспечивают принцип самоподобия и плотнейшую упаковку. Начиная с нулевой точки распространение каркаса в пространстве осуществляется присоединением к фиксированным узлам новых биоединиц. Реализация связей происходит по прямым на К-поверхности с периодом, значительно превышающим период каркаса. Это обеспечивает формирование модуля дифференцировки путем оконтурирования замкнутого и еще не заполненного биоединицами каркаса. Последующее заполнение вакантных узлов каркаса идет без существенных увеличений его линейных размеров. Алгоритм для компьютерного построения модельных модулей дифференцировки с помощью групп переноса
где Литература
|
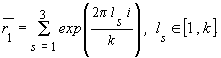
| (c) АСФ России, 2001 |